 Aston-Jones Lab Research Programs
Aston-Jones Lab Research Programs
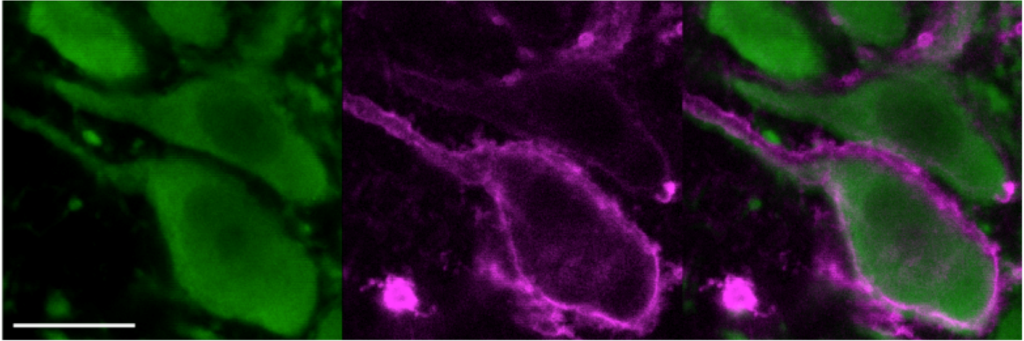
Role of locus coeruleus-noradrenergic system in behavior and cognitive processes. I have conducted studies of the roles of brain modulatory systems in cognitive function throughout my career. We pioneered unit recordings from locus coeruleus (LC) noradrenergic neurons in behaving rats and monkeys, and currently use tetrodes and multi-electrode arrays to record from LC and cortical neurons in rats performing cognitive tasks. We developed prominent theories of LC function, culminating in the Adaptive Gain Theory (Aston-Jones and Cohen, Ann. Rev. Neurosci. 2005). This theory describes a role for the LC system in attention and cognitive flexibility, and in several disorders of cognitive function.
- Aston-Jones, G. and Bloom, F. Activity of norepinephrine-containing locus coeruleus neurons in behaving rats anticipates fluctuations in the sleep-waking cycle. J. Neurosci. 1: 876-886 (1981). [>1,300 citations].
- Aston-Jones, G., Ennis, M., Pieribone, V., Nickell, W. and Shipley, M., The nucleus locus coeruleus: restricted afferent control of a broad efferent trajectory, Science 234: 734-737 (1986). [>500 citations].
- Usher, M., Cohen, J.D., Servan-Schreiber, D., Rajkowski, J., and Aston-Jones, G., The role of locus coeruleus in the regulation of cognitive performance. Science 283: 549-554 (1999). [>400 citations].
- Aston-Jones, G and Cohen, J. D., An integrative theory of locus coeruleus-norepinephrine function: adaptive gain and optimal performance. Ann. Rev. Neurosci. 28: 403-450 (2005). [>900 citations].
- Keren, N., Taheri, S., Vazey, E., Morgan, P., Granholm, A.-C., Aston-Jones, G and Eckert, M. Histologic validation of locus coeruleus MRI contrast in post-mortem tissue. Histologic validation of locus coeruleus MRI contrast in post-mortem tissue. Neuroimage 113, 235–245 (2015). http://doi.org/10.1016/j.neuroimage.2015.03.020. PMCID: In Progress.
Role of orexin neurons in reward processing and addiction. We were first to discover the role of orexin/hypocretin in drug seeking (Harris et al., 2005), and this finding spawned a large and growing body of work into the role of this neuropeptide system in reward and addiction. Our findings led us to propose the functional dichotomy hypothesis, where lateral orexin neurons are involved in reward seeking and medial orexin neurons play a prominent role in arousal and stress (Harris & Aston-Jones, 2006). We have extended our studies of orexin’s role in drug abuse to also document an important role in conditioned food seeking. We recently published an extensive review and perspectives paper, that proposes a primary role for orexins in motivational activation (Mahler et al., 2014).
- Harris, G., Wimmer, M. and Aston-Jones, G., A role for lateral hypothalamic orexin neurons in reward seeking. Nature 437: 556-559 (2005). [>600 citations].
- Harris, G. and Aston-Jones, G., Arousal and reward: A dichotomy in orexin function. Trends Neurosci. 29: 571-577 (2006).
- Sartor, G and Aston-Jones, G., A septal-hypothalamic pathway drives orexin neurons and cocaine seeking. J. Neurosci. 32: 4623-4631 (2012).
- Mahler, S.V., Smith, R.A., Moorman, D.E., James, M. and Aston-Jones, G., Motivational activation: A unifying hypothesis for the orexin/hypocretin system. Nature Neurosci 17: 1298-1303 (2014).
Use of designer receptors to test causal roles of specific brain circuits in behavioral functions. Our lab has implemented DREADD (Designer Receptors Exclusively Activated by Designer Drugs) technology in behavioral and electrophysiological studies. Genes for these receptors are expressed using viral vector microinjections in vivo, resulting in strong DREADD expression in locus coeruleus noradrenergic (LC-NE) neurons, ventral tegmental area dopaminergic (VTA-DA) neurons (in TH-Cre rats), orexin neurons, CAM Kinase neurons, and other neurons. Expression was often directed in a cell-type specific manner using promoters or TH-Cre rats. Results with these methods have revealed an important role for LC-NE neurons in arousal and general anesthesia, and in cognitive dysfunctions in an animal model of Down Syndrome/Alzheimer’s Disease, and for VTA-DA neurons in cued reinstatement of cocaine seeking.
- Vazey, E. and Aston-Jones, G., New tricks for old dogmas: Optogenetic and pharmacosynthetic insights for Parkinson’s disease. Brain Res. 1511: 153–163 (2013). PMCID: PMC3654066
- Vazey, E.M. and Aston-Jones, G., Designer receptor manipulations reveal role of locus coeruleus noradrenergic system in isoflurane general anesthesia. Proc. Nat. Acad. Sci. 111(10): 3859-3864 (2014); doi: 10.1073/pnas.1310025111. PMCID: PMC3956184
- Mahler, S.V., Vazey, E.M., Beckley, J.T., Kaufling, J., Keistler, C.R., McGlinchey, E.M., Wilson, S.P., Deisseroth, K., Woodward, J.J. and Aston-Jones, G., Designer Receptors Show Role for Ventral Pallidum Input to Ventral Tegmental Area in Cocaine Seeking. Nature Neurosci. 17: 577-585 (2014); doi: 10.1038/nn.3664 PMCID: PMC3973180
- Fortress, A., Hamlett, E.D., Vazey, E., Aston-Jones, G., Cass, W., Boger, H.A. and Granholm, A.-C., Designer Receptors Enhance Memory in a Mouse Model of Down Syndrome. J. Neurosci. 35(4):1343-53 (2015). PMCID: In Progress
Development of a novel behavioral economics approach to measure motivation. We recently developed a method for generating and quantitatively analyzing demand curves from within-session behavioral economics tasks, using a progressively increased cost of reward. This method generates normalized measures of demand at low effort (Q0) and motivation (alpha). We have applied versions of this method to self-administration of cocaine, methamphetamine, remiphentanil, all of which generated demand curves that fit closely to exponential demand equations and thus generated accurate estimates of Q0 and alpha. This approach revealed that alpha (motivation) for cocaine predicts subsequent relapse behavior, punished responding for drug, and efficacy of treatment with oxytocin in a rat self-administration model. These findings, and the fact such economic demand curve methods have been applied successfully in humans, lead us to propose that alpha is a valuable biomarker for addiction liability in people.
- Bentzley, B., Fender, K. and Aston-Jones, G., The behavioral economics of drug self-administration: A review and new analytical approach for within-session procedures. Psychopharmacology 226:113–125 (2012). PMCID: PMC3572328
- Bentzley, B., Jhou, T. and Aston-Jones, G., Economic demand predicts addiction-like behavior and therapeutic efficacy of oxytocin in the rat. Proc. Nat. Acad. Sci. 111 (32): 11822-11827 (2014); doi/10.1073/pnas.1406324111. [PNAS press release] PMCID: PMC4136574
- Bentzley, B. and Aston-Jones., Economic demand for cocaine is attenuated by an orexin?/?hypocretin-1 receptor antagonist and predicts reductions in cue-induced reinstatement of cocaine seeking in the rat. Europ. J. Neurosci. 41(9):1149-56 (2015). PMCID: In progress
Afferent regulation of ventral tegmental area. Our lab conducted anatomical, neurophysiological and behavioral studies into afferent regulation of neurons in the ventral tegmental area. These studies revealed novel and topographically specific regulation by the bed nucleus of the stria terminalis, locus coeruleus and medullary noradrenergic cell groups, ventral pallidum, and several other areas. These results also revealed circuits that the VTA participate in to regulate motivated and addiction-related behaviors.
- Georges, F. and Aston-Jones, G., Activation of ventral tegmental area cells by the bed nucleus of the stria terminalis: a novel glutamate input to midbrain dopamine neurons. J. Neurosci. 22: 5173-5187 (2002).
- Mahler, S.V., Vazey, E.M., Beckley, J.T., Kaufling, J., Keistler, C.R., McGlinchey, E.M., Wilson, S.P., Deisseroth, K., Woodward, J.J. and Aston-Jones, G., Designer Receptors Show Role for Ventral Pallidum Input to Ventral Tegmental Area in Cocaine Seeking. Nature Neurosci. 17: 577-585 (2014). PMCID: PMC3973180
- Moorman, D. and Aston-Jones, G., Orexin/Hypocretin modulates response of ventral tegmental dopamine neurons to prefrontal activation: Diurnal influences. J. Neurosci. 30: 15585-15599 (2010). PMCID: PMC 3004096
- Luo, A.H., Tahsili-Fahadan, P., Wise, R.A., Lupica, C.R. and Aston-Jones, G., Linking context with reward: A functional circuit from CA3 of hippocampus to ventral tegmental area. Science 333: 353-357 (2011). PMCID: PMC3150711
Role of protracted withdrawal in drug reward. We studied brain mechanisms of affective aversion produced by opiate withdrawal. Additional studies established that abstinence-withdrawal from morphine or cocaine produced a persistent anxiety state that increased the rewarding value of subsequent acute drug exposure; we propose that this occurs via negative reinforcement. Other of our studies revealed that this increase in resting anxiety and associated increase in drug reward was mediated through noradrenergic innervation of the bed nucleus of the stria terminalis (BNST), consistent with previous findings linking BNST and NE with anxiety. We also found a novel projection from BNST to VTA, providing a possible circuit connection whereby protracted withdrawal effects could be linked to acute reward.
- Delfs, J., Zhu, Y., Druhan, J. and Aston-Jones, G., Noradrenaline in ventral forebrain is critical for opiate withdrawal-induced aversion. Nature 403: 430-434 (2000).
- Harris, G. and Aston-Jones, G., Altered motivation and learning following opiate withdrawal: Evidence for prolonged dysregulation of reward processing. Neuropsychopharmacology 28: 865-871 (2003).
- Aston-Jones, G. and Harris, G., Brain substrates for increased drug seeking during protracted withdrawal. Neuropharmacology 47S1: 167-179 (2004).
- Harris, G. and Aston-Jones, G., Activation in extended amygdala corresponds to altered hedonic processing during protracted morphine withdrawal. Behav. Brain Res. 176 : 251 - 258 (2007).
Publications are available in MyBibliography at http://www.ncbi.nlm.nih.gov/sites/myncbi/gary.aston-jones.1/bibliography/40466698/public/?sort=date&direction=ascending.